1. Algorithms for Dirichlet tessellation of spatial points are developed and
implemented on personal computer. Up to 3000 tessellations of points in an area of any
rectangular dimensions can be scaled appropriately and viewed on computer screen or output
to laser printer.
2. The program also calculates Dirichlet cell areas and their coefficient of variation
(CV) as well as the average nearest neighbour distance between points.
3. Simulations revealed the polynomial relationship between the CV and the
minimum spacing between points. The relationship is used to predict the percentage of
maximum spacing that is exhibited by a population. This value times the maximum spacing
distance possible between objects in an area (hexagonal arrangement) yields the minimum
allowed distance (MAD) that is characteristic of individuals of some territorial or
`inhibitive' species.
4. The program and relationship were used to analyze the spatial attack patterns
of the bark beetles, Dendroctonus brevicomis LeConte, Tomicus piniperda (L.),
and Pityogenes chalcographus (L.) and determine their MAD's. All three species
exhibited spacing between attack sites, in agreement with known behavioural mechanisms that
are proposed for avoiding intraspecific competition for food resources.
The distribution and abundance of organisms can be represented by spatial points in a plane.
A Dirichlet tessellation surrounds a point as a planar polygon in which all regions are closer
to the point than to any other points. This tessellation was proposed in 1850 by Dirichlet
(Upton & Fingleton 1985) and a formal mathematical definition is given by Green & Sibson
(1978). The latter authors state that "the Dirichlet tessellation is one of the most fundamental
and useful constructs determined by an irregular lattice." The Dirichlet tessellation cell, also
known as Voronoi or Thiessen polygons, has been reinvented several times and is useful to
research in many scientific fields (Rogers 1964; Mead 1971; Rhynsburger 1973; Upton &
Fingleton 1985; David 1988; Galitsky 1990).
Dirichlet tessellations can be thought of as representing the areas of territorial animals,
allelochemic-producing plants, or the packing of cells in a tissue. For example, two adjacent
points, representing competitive animals of equal strength, bisect the planar area between
them as well as with any other nearby animals. In general, competitors that are farther away
from an organism will be less likely to interfere spatially unless there are no other organisms
in between that can contest the areas. Thus, the areas of Dirichlet tessellations should
coincide generally with the areas of the territorial or competitive ranges of the organisms
(Tanemura & Hasegawa 1980; Kenkel, Hoskins & Hoskins 1989a, b).
The first computer algorithm for drawing Dirichlet cells was offered by Green & Sibson
(1978). These authors developed a program in ANSI FORTRAN for use on mainframe
computers that has been subsequently utilized in spatial statistics textbooks (Ripley 1981;
Diggle 1983; Upton & Fingleton 1985). Another algorithm has been described in Russian and
programmed in FORTRAN IV (Galitsky 1990). Dirichlet (Voronoi) cells have been delineated
by algorithms that use Delaunay triangles and circumscribing circles on a HITAC M-180
computer (Tanemura & Hasegawa 1980) or elimination of intersecting circles according to a
set of rules (Honda 1978). Unfortunately the above algorithms are described in only general
terms, or in the program code, so they are generally difficult to use. Recently, the commercial
statistical software, SYSTAT 5.0 (Wilkinson 1990), has offered graphical plotting of Voronoi
polygons. Wilkinson (1990) says the algorithms of Green & Sibson (1978) were not used, but
no references or algorithms are presented. Since none of the previous methods nor SYSTAT
calculate areas and variance of Dirichlet cells, my objectives were both to develop algorithms
for drawing tessellations with personal computers and to calculate cell areas. These general
procedures could then be used specifically to analyze the spatial distributions of bark beetle
`attacks' on their host trees. Statistical regression using the coefficient of variation of cell areas
revealed a new method for analyzing spatial point distributions. In addition, these analyses
offer a second way of determining species-specific spacing distances, termed the minimum
allowed distance (MAD) as proposed earlier (Byers 1984).
Methods
A computer program, coded in BASIC, implementing Dirichlet tessellation algorithms was
developed for personal computer that allows x- and y-coordinates of spatial data
in any units to be entered into a file for later retrieval. The data files are compatible with
another program for drawing contour maps of point densities (Byers 1991a). Alternatively, one
can generate x,y coordinates at random, with or without a degree of minimum spacing.
For a given rectangular area (AREA) containing N points, the maximum distance
possible to space apart points is given by 1.0746/SQRT(N/AREA) [where SQRT is square root], which means the points
are in a perfect hexagonal arrangement (Clark & Evans 1954). Thus, an input value of no
more than about 70 percent of the maximum distance should be attempted since the computer
otherwise may not find locations for all the points due to constraints from the initial selections.
The algorithms for spacing points have been described earlier (Fig. 1 in Byers 1984).
Once the x,y coordinates of the points are entered, an inner border area should be
chosen in order to avoid tessellating points on the periphery. Points near the edges of the
area are not surrounded by other points so the tessellation outline would be altered by the
boundary of the area. It was found empirically that for randomly distributed points the use of
a border width of at least 1.5 times the distance expected for the fourth nearest
neighbour (1.0937/SQRT(N/AREA), Thompson 1956) gave good results. The distance used, however,
is arbitrary and can be adjusted.
Dirichlet tessellation algorithms
The program draws tessellations about each point within the inner border area, although all
points including those in the peripheral area are considered. Real coordinates and dimensions
are scaled on the monitor screen as well as on laser printers. The algorithms are described
in six steps:
(1) The first step is to find the nearest neighbours which might affect the Dirichlet cell outline.
Coordinates of all points (N) are scanned to count the number of points contained
within a `box' centered about the point in question. The size of the initial box is smaller than
that expected to hold p neighbours (equal to N-1 or 35 points, whichever is
smaller) if they were distributed at random (0.2(SQRT[(AREA/N)P]/2)). However, if less than
p points (hereafter 35 points) fall within the box, then the box is successively enlarged
by 10 percent until the required points are obtained. The actual distances from the `center'
(xc, yc) point to its neighbours (xn,
yn) in the box are then computed, dn = SQRT ((xc - xn)2 + (yc - xn)2), and the 35
lowest distances are sorted with an exchange sort algorithm.
(2) The next step is to calculate the equations of the lines that are perpendicular bisectors
between the center point and each of its 35 neighbours (as well as the four boundary lines).
The 35 resulting equations have the form anx + bny +
cn= 0, where an = 2xc -
2xn,
bn = 2yc - 2xn, and cn
= xnxn - xcxc +
ynyn - ycyc . In Fig. 1, a simplified case
is shown where a center point is surrounded by six neighbours with six perpendicular bisector
lines (solid lines).
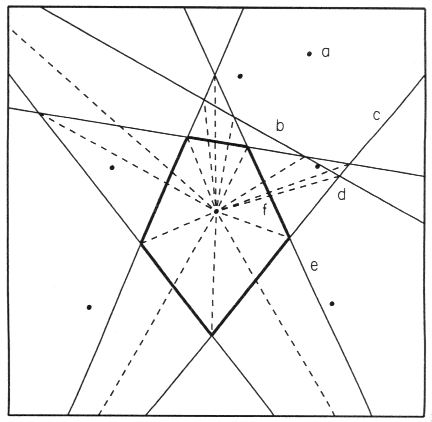
Fig. 1. Dirichlet tessellation (irregular pentagon) composed of perpendicular bisectors (solid
lines) between center point and six surrounding points. Dashed lines connect the center point
with the 15 intersection coordinates of the bisector lines. Bisector line b between the
center and point a is included in the calculation but in this case was not necessary for
the drawing of the Dirichlet cell (see text for more details).
(3) The above 35 equations plus the four boundary equations are then compared to each
other non-redundantly to obtain a total of SUM from j=1 to n-1 of j, or 741 possible intersection coordinates.
The xj,yjcoordinates of the intersection of two such equations,
a1x1+ b1y1+ c1 =
0 and a2x2+ b2y2+ c2=
0, are xj = (-c1b1 +
c2b2)/(a1b2 -
a2b1) and yj = (-a1c2
+ a2c1)/(a1b2 -
a2b1).
(4) The program then calculates the equations of the 741 lines between the center point
(xc,yc) and each of the intersection coordinates
(xj,yj), where aj = yj -
yc, bj = -(xj - xc),
and cj = -ycbj - xcaj.
In Fig. 1 these lines are represented by the 15 dashed lines (lines to the boundary
intersections are not shown).
(5) The perpendicular bisector lines, found in (2) above, are compared to each of the dashed
lines (Fig. 1), found in (4) above, to see if any intersections occur (x,y; method as in
part 3 above) but only in the segment from the center point to and including the intersection
point of the two respective bisectors (xj,yj; from part 3).
Thus, if the x-coordinate is greater or smaller than both xc and
xj or the y-coordinate is greater or smaller than both
yc and yj then no intersection can occur. If the number of
intersections is more than two (both intersecting bisector lines intersect with a dashed line)
then the intersection point found in (3) can not be one of the legitimate vertices of the Dirichlet
cell. This can be seen in one case in Fig. 1 where the bisector line b, between point
a and the center, intersects the bisector line c at d (a dashed line
connects d to the center) but bisector line e intersects the dashed segment at
f, thus invalidating the intersection coordinates at d as a vertex of the Dirichlet
cell.
(6) The final step takes the coordinates of the true vertices of the Dirichlet cell and sorts them
in ascending order by angular direction from the center point. This must be done since it is
not yet known what the correct order of drawing is between vertices. The general method for
obtaining polar coordinates uses cos ŕ = x/r and the appropriate quadrant (Batschelet 1979,
p. 121).
Coefficient of variation of Dirichlet cell areas
The Dirichlet cell area (A) is calculated by means of summing the areas of the
triangles constructed from the center point and the vertices
(xi,yi):
A = SUM from i=1 to k of ABS(0.5(xc(yi - yi+1) + xi(yi+1 - yc) + xi+1(yc - yi)))
where k = number of vertices and xk+1,yk+1 are equal to x1,y1.
The mean and standard deviation (SD), SQRT [N SUM A2n - (SUM An)2)/(N(N - 1))], of the
cell areas are used to calculate the coefficient of variation, CV = SD/mean x
100. Computer simulations were carried out to determine the relationship between the
CV and the degree of uniformity in spacing apart of points. A square area of 447.21
units on a side had 250 points placed within it to obtain a density of 0.00125 per unit area.
Points in areas were increasingly spaced apart at distances from 0 (random) to 70 percent of
the maximum possible spacing (30.39 units) in increments of 10 percent. A total of 32 point
sets, each of 250 points, at each of the spacing constraints were simulated. The algorithms
for spacing have been reported earlier (Byers 1984) and comprise an inhibition model where
points are sequentially placed at random unless they are closer to an established point than
the minimum allowed distance (MAD). The inner border width used for the points was
2.5 times the expected fourth nearest neighbour distance (random distribution), 77.36 units.
The CV of the cell areas will be distorted if tessellations are attempted on points
near the edges of the area. Thus a border area must be chosen that is a compromise
between reducing the "edge effects" and having sufficient points remaining for statistical
analysis. As mentioned above, it was found that a distance of 1.5 times the expected fourth
nearest neighbour distance gave adequate results. The border can be somewhat larger if
more points are available (> 100). The effect of changing the border width on the
CV and the estimated percentage of the maximum point spacing was investigated by
using increasingly larger widths during tessellations of natural bark beetle attack patterns
(described below).
Analysis of bark beetle attack patterns
The patterns of attack entrances of the bark beetles Dendroctonus brevicomis
LeConte on ponderosa pine, Pinus ponderosa Doug. ex. Laws., Pityogenes
chalcographus (L.) on Norway spruce, Picea abies (L.) Karst., and Tomicus
piniperda (L.) on Scots pine, Pinus sylvestris L., were recorded from bark samples
in the field. This was done by overlaying a plastic sheet on each of the bark areas and
marking the plastic with ink. The marks were then measured for x and y
coordinates. An average nearest neighbour distance analysis was done on the attack
distributions (Clark & Evans 1954; Thompson 1956) with a previously described computer
program (Byers 1984). A minimum allowed distance analysis (MAD) using six
simulation runs of 300 points at each of eight spacing steps also was performed on each of
the attack densities (Byers 1984). A simulated placement of 179 attacks at random was done
by the computer program for comparison with the natural attacks of P. chalcographus
above. Dirichlet tessellations were then done on the point patterns to evaluate the program
as well as the attack distributions.
Results
Coefficient of variation of Dirichlet cell areas
Simulated placements of 250 points at a density of 0.00125 points per unit area revealed the
relationship (Fig. 2) between the minimum distance of separation between points and the
coefficient of Dirichlet cell area variation (CV).
Fig. 2. See correction. Relationship between the coefficient of variation (CV) of Dirichlet cell areas and
the minimum spacing between points in any area expressed as the percentage of the
maximum possible point spacing if the points were hexagonally arranged. The vertical lines
represent 95 percent confidence limits (n = 32 simulations of 250 points each per
spacing increment; about 100 tessellations per simulation were analyzed).
The minimum separation distance
between otherwise randomly placed points was increased in increments of ten percent of the
maximum hexagonal spacing distance possible at this density (i.e. 30.39 units, Clark & Evans
1954). It was found that the CV of cell areas was about 55.6 % regardless of the
density or number of points when simulating random point distributions (tests up to 3000
points). At the maximum point spacing possible it is intuitive that the CV of the areas
would be zero since all the cells are perfect hexagons of identical size. This `point' can not
be found by simulation, although the theoretical value was used with the simulated points to
find the best fitting cubic equation (Fig. 2).
Several indexes of dispersion have been proposed to describe the degree of uniformity
or aggregation among spatial points representing organisms (Clark & Evans 1954; Pielou
1959; Morisita 1965; Lloyd 1967; Goodall & West 1979). One of the most widely used is the
R index of Clark & Evans (1954) which is the ratio of the observed nearest neighbour
distance to the expected nearest neighbour distance when points are distributed randomly.
A value greater than 1 indicates that points are more uniformly spaced than at random. It is
significant that this ratio is independent of density. In Fig. 2, the CV of Dirichlet cells
is also independent of density. Thus, one could tessellate a spatial point pattern and
determine the CV and then use the cubic regression equation (Fig. 2) to obtain a value
for the percentage of the maximum spacing that organisms exhibit. Solving algebraically for
X in polynomial equations is not possible but must be done by `binary successive
approximation' until a value is found that gives the known Y (CV). This method
has been incorporated into the program. For an algebraic solution, the best quadratic
regression is Y = aX2 + bX + c where a = -
.002386, b = -.354, and c = 57.9; and X = (-b - SQRT (b2 - 4a(c-Y))) / 2a.
Earlier I proposed a method called the minimum allowed distance (MAD) which
purports to find the preferred or instinctive minimum distance that individuals will space
themselves apart from others (Byers 1984). Beyond this species-specific distance individuals
are free to colonize sites at random. The method relies on construction of a quadratic
regression curve from computer simulations of increasing minimum allowed distances of
separation and the resulting average nearest neighbour distances obtained at a density
corresponding to natural spatial data. The observed nearest neighbour distance for the natural
data is then used to solve the quadratic equation to obtain the MAD for the species.
This distance is independent of density since it is based on a behavioural distance that is
relatively constant regardless of density.
It now is apparent that one may also find the MAD, assuming one exists, from the
CV of the Dirichlet areas. Cells with a lower CV than expected indicate that the
points are more uniformly spaced, and from the relationship in Fig. 2 one may find the
percentage of the maximum point spacing at a particular density. This percentage multiplied
by the maximum point spacing distance is equal to the MAD. The results of the two
methods will be compared subsequently for several examples of the bark beetle attacks.
Attack spacing of Ips typographus on Norway Spruce
Analysis of bark beetle attack patterns
The CV of the Dirichlet cells becomes incorrectly large when the border within which
tessellations are drawn is made too narrow. However, an increase in the width of the border
after a certain amount does not appreciably effect the magnitude of the CV and the
estimate of the percentage of maximum spacing (Fig. 3).
Fig. 3. Effect of border width on the estimated percentage of maximum spacing value
(obtained from the cubic regression in Fig. 2) for the attack patterns of the bark beetles
Pityogenes chalcographus (P. c.), Tomicus piniperda (T. p.), and
Dendroctonus brevicomis (D. b.) and for a simulated pattern of randomly placed
points (R). Vertical lines represent 95 percent confidence limits. The numbers along
the dashed line are the number of attacks that were both tessellated and within the area inside
the border.
The estimate of the percentage of
maximum spacing was relatively constant (Fig. 3) for different sized areas (and numbers of
Dirichlet cells) for the attacks of Pityogenes chalcographus and Tomicus
piniperda as well as the random point distribution, but not for the attacks of
Dendroctonus brevicomis (data from Figs. 4-6). This indicates that the samples of the
former two species and the random pattern are consistent at all scales while the data for D.
brevicomis is more variable.
The pattern of attacks of D. brevicomis can be seen in Fig. 4.
Fig. 4. Dirichlet cell tessellations of 35 attacks of Dendroctonus brevicomis inside a
border width of 1.5 times the expected fourth nearest neighbor distance in an area of 90 x 45
cm (n=97). The average cell area is 33.9 cm2 ± 5.4 (95% confidence limits)
and the coefficient of variation (CV) is 48%. The percentage of maximum spacing was
28.1 (0-39.3, 95% confidence interval, Fig. 2).
The average Dirichlet
cell area was 33.9± 5.4 cm2 (± 95% confidence limits, C.L.). The
coefficient of variation in the cell areas was 48% (41.5-57.1%, 95% confidence interval, C.I.)
which yields a spacing value of 28.1% (0-39.3%, 95% C.I.), of the maximum possible spacing
as estimated from cubic regression (Fig. 2). The estimated MAD using the Dirichlet
CV method is then 0.281 x [1.0746/SQRT(97/(90x45))] = 1.95 cm (0-2.73 cm, 95% C.I.). The observed average nearest
neighbour distance was 3.63± 0.39 cm (95% C.L.) and the expected corresponding
distance if points were random is 3.23± 0.34 which gives a R = 1.12, indicating
a significant degree of spacing (P=0.02, Clark & Evans 1954). The alternative analysis using
the nearest neighbour distances and simulation of spaced points (Byers 1984) gave a
MAD = 1.67 cm (0.38-2.56 cm, 95% C.I.). The two methods give slightly different
estimates of the MAD possibly because the spatial distribution of attacks was not
consistent at different border widths (Fig. 3).
The average Dirichlet cell area for Tomicus piniperda (Fig. 5) was 33.4± 2.4
cm2 with a CV = 24.5% (22.8-26.4%, C.I.) and a maximum spacing
percentage of 62.8 (60.3-64.9%, C.I.).
Fig. 5. Dirichlet cell tessellations of 44 attacks of Tomicus piniperda inside a border
width of 1.5 times the expected fourth nearest neighbor distance in an area of 86.5 x 50 cm
(n=108). The average cell area is 33.4 cm2 ± 2.4 (95% confidence limits) and
the coefficient of variation (CV) is 24.5%. The percentage of maximum spacing was
62.8 (60.3-64.9, 95% confidence interval, Fig. 2).
The MAD was thus estimated to be 4.27 cm
(4.10-4.42, C.I.). The average nearest neighbour distance was 4.62± 0.19 cm (C.L.), the
expected distance was 3.16± 0.31, giving a R = 1.46, indicating significant
spacing (P < 0.001). The nearest neighbour simulation analysis estimated the MAD
to be 3.71 cm (3.4-4 cm, C.I.).
Dirichlet tessellations of Pityogenes chalcographus attacks (Fig. 6a) produced cells
with an average area of 6.87± 0.44 cm2 and a CV = 30.28% (28.4-
32.4%, C.I.) yielding a maximum spacing percentage of 55% (52.2-57.5%, C.I.).
Fig. 6 (a). Dirichlet cell tessellations of 84 attacks of Pityogenes chalcographus inside
a border width of 1.5 times the expected fourth nearest neighbor distance in an area of 30 x
45 cm (n=179). The average cell area is 6.87 cm2 ± .44 (95% confidence
limits) and the coefficient of variation (CV) is 30.3%. The percentage of maximum
spacing was 55 (52.2-57.5, 95% confidence interval, Fig. 2); (b) Dirichlet cell tessellations of
91 points placed at random inside a border width of 1.5 times the expected fourth nearest
neighbor distance in an area of 30 x 45 cm (n=179). The average cell area is 6.74
cm2 ± .81 (95% confidence limits) and the coefficient of variation (CV)
is 58.8%. The percentage of maximum spacing was 0 (0-18.2, 95% confidence interval, Fig.
2).
The
MAD was thus calculated to be 1.62 cm (1.54-1.7, C.I.). The average nearest
neighbour distance was 1.99± 0.1 cm (C.L.), the expected distance was 1.37± 0.11,
giving a R = 1.45, indicating significant spacing (P < 0.001). The nearest neighbour
simulation analysis estimated the MAD to be 1.58 cm (1.42-1.75 cm, C.I.). The
estimates of the two methods are very close. In comparison, the random distribution at the
same point density (Fig. 6b) gave a CV = 58.8% which, as expected, was not different
from random distributions (Fig. 2 at 0). The average nearest neighbour distance was 1.47 cm
(greater than the expected distance) and gave a R = 1.07, but this was not statistically
significant from R = 1 for a random pattern (P=0.08).
Discussion
The Dirichlet cell was first proposed in 1850 but has been rediscovered several times and
given names such as Voronoi polygons, 1909, Thiessen polygons, 1911, Wigner-Seitz cells,
1933, the cell model, 1953, and the S-mosaic, 1977, (Upton & Fingleton 1985). For a
theoretical Poisson forest, the expected number of sides of the Dirichlet cell is 6, the expected
area is , and the expected perimeter length is , where d is the density of
`trees' (Meijering 1953). Matérn (1979) calculates the expected length of border areas
between Dirichlet cell mosaics of two species, if the distributions of the species are
random.
Applications of the Dirichlet cell in plant ecology and forestry have been discussed with
regard to interplant competition and prediction of growth for individual trees (Brown 1965;
Mead 1971; Cormack 1979; Kenkel, Hoskins & Hoskins 1989a, b; Welden, Slauson & Ward
1990). Dirichlet polygons describe territories of pectoral sandpipers, Calidris melanotos,
male mouthbreeder fish, Tilapia mossambica, and nest areas of Royal terns, Sterna
m. maxima (Grant 1968; Barlow 1974; Buckley & Buckley 1977). The cellular patterns of
coenobial green algae, Pediastrum boryanum, as well as cultured epithelial cells of
chicks (retinal and lung), rat (intestine) and mudpuppy (gallbladder) show Dirichlet packing
(Honda 1978).
Boots & Murdoch (1983) used Monte Carlo procedures (programmed in FORTRAN IV) to
investigate the properties of Dirichlet tessellation of random points. Their program, however,
is not generally useful to ecologists since "there is no input to the program". It is not known
if the algorithms used in the present study are as efficient as those of Green & Sibson (1978),
Honda (1978) or Tanemura & Hasegawa (1980). However, the use of a math-coprocessor
allows drawing of 500 cells within a few minutes by personal computer (computation time is
similar to that for SYSTAT). The computation cost of the algorithm of Green & Sibson (1978)
increases roughly as n1.5 (Diggle 1983), while the computational time for the
algorithm presented here increases as n1.36 (geometric regression, n=7).
Dirichlet polygons can represent the competitive interactions of a colony of bark beetles
packed onto the bark surface. Most temperate bark beetles (Coleoptera: Scolytidae), including
Dendroctonus brevicomis, Tomicus piniperda, and Pityogenes chalcographus
attack the outer bark and bore into the thin layer of phloem/cambium covering the woody
xylem tissue of trees. The beetles construct a two-dimensional system of tunnels or galleries
within the layer where they feed and reproduce. The thickness of the layers is similar to that
of a beetle so competition is expected to be severe for this limited food resource. In fact,
reports of intraspecific competition in several species in the genera Ips,
Dendroctonus, Scolytus and Tomicus have shown that brood output per
female decreases at higher attack densities (Miller & Keen 1960; McMullen & Atkins 1961;
Eidmann & Nuorteva 1968; Ogibin 1973; Beaver 1974; Mayyasi et al. 1976; Wagner et al.
1981; Light, Birch & Paine 1983; Anderbrant, Schlyter & Birgersson 1985).
Bark beetles can minimize potential competition by avoiding areas releasing pheromone
components that indicate higher densities of established individuals (Byers et al. 1988; Byers
1989). Another mechanism that may require little time and energy expenditure to gain large
advantages in reproductive success is to avoid boring too closely to established attack holes
and their galleries. Several bark beetle species, including Tomicus piniperda, are
known to space their attacks (Nilssen 1978; Byers 1984) and this is evident also for
Dendroctonus brevicomis and Pityogenes chalcographus (Figs 3, 4 and
6a).
Dendroctonus brevicomis is the most important pest bark beetle of forests in
California (Miller & Keen 1960). The female initiates the attack and bores a sinusoidal tunnel
under the bark in the phloem layer. She produces a pheromone component, exo-
brevicomin, which attracts primarily males (Silverstein et al. 1968; Byers 1989). The male
arrives and joins a female in her gallery and he releases a second attractive pheromone
component, frontalin, which is synergistic with exo-brevicomin (Kinzer et al. 1969;
Wood et al. 1976). This causes beetles to aggregate en masse and overcome the tree's
resistance, resulting in its death and successful reproduction by the beetle. Over-crowding
would result if not for several mechanisms, only partly understood, to avoid severe competition
(Byers 1989).
Both sexes produce trans-verbenol which at close-range inhibits the female sex from
entering holes releasing attractive pheromone components (Byers 1983). Verbenone, is
produced by males and this compound as well as trans-verbenol inhibits both sexes
from flying to sources releasing attractive pheromone (Bedard et al. 1980; Byers et al. 1984).
Still another compound, ipsdienol, is produced in small amounts by males and inhibits both
sexes (Byers et al. 1984). Thus, these compounds may function together to limit the overall
attack density as well as close-range spacing.
Another mechanism that has been postulated to regulate density of attack is acoustic
stridulation. Compared to males, females stridulate very weakly and it has been reported that
females increased their chirping rate when other stridulating females were boring holes in the
vicinity (Rudinsky & Michael 1973). However, male chirps can be heard from the onset of
colonization, and for several days, from even a meter or more away by the human ear (Byers
et al. 1984). Vibrations from the male stridulation could possibly be felt by walking females
which would then decide to leave the area. Alternatively, males within holes with females may
stridulate to warn walking males not to attempt entry into their tunnels. The function of
stridulation is poorly understood.
Nilssen (1978) used nearest neighbour analysis to show that the pattern of attacks of
Tomicus piniperda was more uniform than a random pattern. Presumably the spacing
of attacks is to avoid competition between larvae as has been shown in simulation models
(DeJong & Saarenmaa 1985). The MAD of from 3.7 to 4.3 cm indicates that T.
piniperda spaces further apart than either D. brevicomis (1.7-2 cm) above or the
European spruce bark beetle, Ips typographus (2.5 cm, Byers 1984).
T. piniperda, although a pest of Scots pine in Europe, does not produce an
aggregation pheromone, as do most bark beetles that aggregate on trees, but instead is
attracted to host volatiles. A combination of the monoterpenes ŕ-pinene, 3-carene, and
terpinolene emanating from wound oleoresin of storm-damaged trees serves in a mechanism
for recognition of the host as well as its susceptibility to attack (Byers et al. 1985). Both sexes
contain a small amount of verbenone in their hindguts (Lanne et al. 1987) which is probably
released with the faecal pellets as are pheromone components in other bark beetle species
(Byers 1989). Verbenone, at a range of a few mm to cm, could inhibit beetles from boring
nearby. Beetle infested logs of Scots pine increasingly released verbenone with time while a
control log released a constant and very low amount (Byers, Lanne & Löfqvist 1989). Release
of verbenone at rates comparable to several infested logs significantly reduced the attraction
of flying beetles to host monoterpenes. This indicates that verbenone may function in spacing
of attacks as well as a cue that the host is now unsuitable for colonization (Byers, Lanne &
Löfqvist 1989). Males of T. piniperda also stridulate audibly so this could be part of a
mechanism for spacing. Stridulation appears to play a role in mate recognition, and
undoubtedly in male-male fighting (Byers 1991b).
In contrast to D. brevicomis and T. piniperda where the female attacks, the
male of Pityogenes chalcographus chooses the attack site on Norway spruce and thus
is responsible for avoiding competition. The males produce two pheromone components,
(E,Z)-(2,4)-methyl decadienoate and chalcogran, which are attractive to both sexes
(Francke et al. 1977, Byers et al. 1988). Several host monoterpenes, including 3-carene, ŕ-
and á-pinene, stimulate entering of artificial holes when the pheromone components are
present, indicating the monoterpenes play a role in host recognition (Byers et al. 1988). There
are no known olfactory inhibitors which might regulate spacing. However, higher release rates
of the attractive pheromone components cause the males, but not apparently the females, to
be less attracted (Byers et al. 1988). This mechanism may inhibit males from boring too
closely to established attacks. Neither sex seems to be able to stridulate. Therefore, a suitable
explanation for spacing in this species is lacking.
Bark structure also has been postulated as enforcing spacing in bark beetles (Safranyik &
Vithayasai 1971). Ponderosa pine has rather deep furrows running longitudinally that branch
in diagonal directions. It was observed that 79 of 97 attacks of D. brevicomis were in
crevices, visible as the longitudinal 'dotted-lines' in Fig 4. The deep crevices in ponderosa pine
allow a more rapid and efficient entry into the phloem, this combined with competition probably
has provided the selection pressure for evolution of the sinusoidal gallery making. The boring
of a gallery parallel to the wood grain, as in many other bark beetles, would be
disadvantageous for D. brevicomis since it would soon encounter attacks of
neighbours. However, a sinusoidal gallery would avoid nearby neighbours immediately above
and below in the crevice.
At the base of Scots pine trees the furrows are more numerous and attacks of T.
piniperda are associated with crevices (all 67 attacks in an area of 64 x 56 cm), but higher
up the trunk the attacks occur more often under bark flakes (as in Fig. 5). It appears that the
density of suitable sites for attack on the bark is higher than the observed attack density. Also,
if the number of bark flakes is limiting then one would expect clumping of attacks - which does
not appear to occur (Fig. 5). However, in the case of P. chalcographus (Fig. 6a) the
bark of Norway spruce was like that of a fine-grained `sandy' surface so bark irregularities
seem unlikely to have caused the spacing between attacks.
The Dirichlet tessellation program and calculation of the MAD of spacing will allow
many other plant and animal species to be analyzed. The program is available from the
author, please send a formatted disk and mailer for IBM-compatible personal computers.
Acknowledgements
Funding for the work was contributed by the Swedish Forest and Agricultural Research
Agency (SJFR). I thank my colleagues Olle Anderbrant and Fredrik Schlyter in the Pheromone
Research Group for review of the manuscript.
John A. Byers
Department of Animal Ecology, Lund University, SE-223 62 Lund, Sweden
Present address:
References
Anderbrant, O., Schlyter, F. & Birgersson, G. (1985). Intraspecific competition affecting
parents and offspring in the bark beetle Ips typographus. Oikos, 45,
89-98.
Batschelet, E. (1979).Introduction to Mathematics for Life Scientists.
Springer-Verlag, New York, USA.
Beaver, R.A. (1974). Intraspecific competition among bark beetle larvae (Coleoptera:
Scolytidae). Journal of Animal Ecology,43, 455-467.
Bedard, W.D., Tilden, P.E., Lindahl, K.Q. Jr., Wood, D.L. & Rauch, P.A. (1980).
Effects of verbenone and trans-verbenol on the response of Dendroctonus
brevicomis to natural and synthetic attractant in the field. Journal of Chemical
Ecology, 6, 997-1013.
Boots, B.N. & Murdoch, D.J. (1983). The spatial arrangement of random Voronoi
polygons. Computers and Geosciences,9, 351-365.
Brown, G.S. (1965). Point density in stems per acre. New Zealand Forestry
Service Research Notes, 38, 1-11.
Buckley, P.A. & Buckley, F.G. (1977). Hexagonal packing of royal tern nests.
The Auk94, 36-43.
Byers, J.A. (1983). Bark beetle conversion of a plant compound to a sex-specific
inhibitor of pheromone attraction. Science, 220, 624-626.
Byers, J.A. (1984). Nearest neighbor analysis and simulation of distribution patterns
indicates an attack spacing mechanism in the bark beetle, Ips typographus
(Coleoptera: Scolytidae). Environmental Entomology, 13, 1191-1200.
Byers, J.A. (1989). Chemical ecology of bark beetles. Experientia,
45, 271-283.
Byers, J.A. (1991a). Grid cell contour mapping of point densities: bark beetle attacks,
fallen pine shoots, and infested trees. Oikos, 62, in press.
Byers, J.A. (1991b). Simulation of the mate-finding behaviour of pine shoot beetles,
Tomicus piniperda. Animal Behaviour, 41, 649-660.
Byers, J.A., Birgersson, G., Löfqvist, J. & Bergström, G. (1988). Synergistic
pheromones and monoterpenes enable aggregation and host recognition by a bark beetle.
Naturwissenschaften, 75, 153-155.
Byers, J.A., Lanne, B.S. & Löfqvist, J. (1989). Host-tree unsuitability recognized
by pine shoot beetles in flight. Experientia, 45, 489-492.
Byers, J.A., Lanne, B.S., Schlyter, F., Löfqvist, J. & Bergström, G. (1985).
Olfactory recognition of host-tree susceptibility by pine shoot beetles.
Naturwissenschaften, 72, 324-326.
Byers, J.A., Wood, D.L., Craig, J. & Hendry, L.B. (1984). Attractive and inhibitory
pheromones produced in the bark beetle, Dendroctonus brevicomis, during host
colonization: Regulation of inter- and intraspecific competition. Journal of
Chemical Ecology, 10, 861-877.
Clark, P.J. & Evans, F.C. (1954). Distance to nearest neighbor as a measure of
spatial relationships in populations. Ecology, 35, 445-453.
Cormack, R.M. (1979). Spatial aspects of competition between individuals. Spatial
and Temporal Analysis in Ecology (Ed. by R.M. Cormack and J.K. Ord), pp. 151-
211. Fairland International Cooperative Publishing House, Maryland, USA.
David, C.W. (1988). Voronoi polyhedra as structure probes in large molecular
systems. VII. Channel identification. Computers and Chemistry, 12, 207-
208.
DeJong, C.M.M. & Saarenmaa, H. (1985). A mechanistic simulation model for the
movement and competition of bark beetle larvae (Coleoptera, Scolytidae).
Ecological Modelling, 27, 109-138.
Diggle, P.J. (1983).Statistical Analysis of Spatial Point Patterns, Academic
Press, New York, U.S.A.
Eidmann, H.H. & Nuorteva, M. (1968). Der Einfluss der Siedlungsdichte und anderer
Faktoren auf die Anzahl Nachkommen von Blastophagus piniperda L. (Coleoptera:
Scolytidae). Annales Entomologica Fennica,34, 135-148.
Francke, W., Heemann, V., Gerken, B., Renwick, J.A.A. & Vité, J.P. (1977). 2-
ethyl-1-6-dioxaspiro[4.4]nonane, principal aggregation pheromone of Pityogenes
chalcographus (L.). Naturwissenschaften, 64, 590-591.
Galitsky, V.V. (1990). Dynamic 2-d model of plant communities. Ecological
Modelling, 50, 95-105.
Goodall, D.W. & West, N.E. (1979). A comparison of techniques for assessing
dispersion patterns. Vegetatio, 40, 15-27.
Grant, P.R. (1968). Polyhedral territories of animals. The American Naturalist,
102, 75-80.
Green, P.J. & Sibson, R. (1978). Computing dirichlet tessellations in the plane.
The Computer Journal, 21, 168-173.
Honda, H. (1978). Description of cellular patterns by Dirichlet domains: the two-
dimensional case. Journal of Theoretical Biology, 72, 523-543.
Kenkel, N.C., Hoskins, J.A. & Hoskins, W.D. (1989a). Edge effects in the use of
area polygons to study competition. Ecology, 70, 272-274.
Kenkel, N.C., Hoskins, J.A. & Hoskins, W.D. (1989b). Local competition in a
naturally established jack pine stand. Canadian Journal of Botany67,
2630-2635.
Kinzer, G.W., Fentiman, A.F. Jr., Page, T.F., Foltz, R.L., Vité, J.P. & Pitman, G.B.
(1969). Bark beetle attractants: Identification, synthesis and field bioassay of a new
compound isolated from Dendroctonus. Nature211:475-476.
Lanne, B.S., Schlyter, F., Byers, J.A., Löfqvist, J., Leufvén, A., Bergström, G., Van
Der Pers, J.N.C., Unelius, R., Baeckström, P. & Norin, T. (1987). Differences in
attraction to semiochemicals present in sympatric pine shoot beetles, Tomicus minor
and T. piniperda. Journal of Chemical Ecology, 13,
1045-1067.
Light, D.M., Birch, M.C. & Paine, T.D. (1983). Laboratory study of intraspecific and
interspecific competition within and between two sympatric bark beetle species, Ips
pini and Ips paraconfusus. Zeitschrift für angewandte Entomologie,
96, 233-241.
Lloyd, M. (1967). Mean crowding. Journal of Animal Ecology, 36,
1-30.
Matérn, B. (1979). The analysis of ecological maps as mosaics. Spatial and
Temporal Analysis in Ecology (Ed. by R.M. Cormack and J.K. Ord). pp. 271-288.
Fairland International Cooperative Publishing House, Maryland, USA.
Mayyasi, A.M., Coulson, R.N., Foltz, J.L., Hain, F.P. & Martin, W.C. (1976).
Functional description of within-tree larval and progeny adult populations of
Dendroctonus frontalis (Coleoptera: Scolytidae). Canadian Entomologist,
108, 363-372.
McMullen, L.H. & Atkins, M.D. (1961). Intraspecific competition as a factor in the
natural control of the Douglas-fir beetle. Forest Science7, 197-203.
Mead, R. (1971). Models for interplant competition in irregularly distributed
populations. Statistical Ecology Volume 2 (Ed. by G.P. Patil, E.C. Pielou & W.E.
Waters). pp. 13-30. Penn State University Press, University Park, USA.
Meijering, J.L. (1953). Interface area, edge length, and number of vertices in crystal
aggregates with random nucleation. Philips Research Reports, 8, 270-
290.
Miller, J.M. & Keen, F.P. (1960). Biology and control of the western pine beetle.
U.S. Department of Agriculture, Miscellaneous Publication No. 800, Washington
D.C., USA.
Morisita, M. (1965). A revision of the methods for estimating population values of the
index of dispersion in the Ië - method. Research Population Ecology,
7, 126-128.
Nilssen, A.C. (1978). Spatial attack pattern of the bark beetle Tomicus
piniperda L. (Col., Scolytidae). Norwegian Journal of Entomology, 25,
171-175.
Ogibin, B.N. (1973). Effect of population density on fertility I. Ips typographus
L. Ekologiya, 5, 66-72.
Pielou, E.C. (1959). The use of point-to-point distances in the study of the pattern
of plant populations. Journal of Ecology, 47, 607-613.
Rhynsburger, D. (1973). Analytic delineation of Thiessen polygons. Geographical
Analysis, 5, 133-144.
Ripley, B.D. (1981).Spatial Statistics, John Wiley & Sons, New York,
U.S.A.
Rogers, C.A. (1964). Packing and Covering. Cambridge Tracts in Mathematics
and Mathematical Physics. No. 54, Cambridge University Press, UK.
Rudinsky, J.A. & Michael, R.R. (1973). Sound production in Scolytidae: stridulation
by female Dendroctonus beetles. Journal of Insect Physiology, 19,
689-705.
Safranyik, L. & Vithayasai, C. (1971). Some characteristics of the spatial
arrangement of attacks by the mountain pine beetle, Dendroctonus ponderosae
(Coleoptera: Scolytidae), on lodgepole pine. Canadian Entomologist, 103,
1607-1625.
Silverstein, R.M., Brownlee, R.G., Bellas, T.E., Wood, D.L. & Browne, L.E. (1968).
Brevicomin: Principal sex attractant in the frass of the female western pine beetle.
Science, 159, 889-891.
Tanemura, M. & Hasegawa, M. (1980). Geometrical models of territory. I. Models
for synchronous and asynchronous settlement of territories. Journal of Theoretical
Biology, 82, 477-496.
Thompson, H.R. (1956). Distribution of distance to nth neighbor in a population of
randomly distributed individuals. Ecology, 37, 391-394.
Upton, G. & Fingleton, B. (1985).Spatial data analysis by example. Volume
1. John Wiley and Sons, New York, USA.
Wagner, T.L., Feldman, R.M., Gagne, J.A., Cover, J.D., Coulson, R.N. & Schoolfield,
R.M. (1981). Factors affecting gallery construction, oviposition, and reemergence of
Dendroctonus frontalis in the laboratory. Annals of the Entomological Society
of America, 74, 255-273.
Welden, C.W., Slauson, W.L. & Ward, R.T. (1990). Spatial pattern and interference
in pinon-jniper woodlands of northwest colorado. Great Basin Naturalist50,
313-319.
Wilkinson, L. (1990).SYGRAPH: The System for Graphics. SYSTAT, Inc.,
Evanston, IL., USA.
Wood, D.L., Browne, L.E., Ewing, B., Lindahl, K., Bedard, W.D., Tilden, P.E., Mori,
K., Pitman, G.B. & Hughes, P.R. (1976). Western pine beetle: specificity among
enantiomers of male and female components of an attractant pheromone. Science,
192, 896-898.

 Download DIRICHLE.EXE for PC
Download DIRICHLE.EXE for PC