Department of Ecology, Animal Ecology, Lund University, SE-223 62 Lund, Sweden
Present address:


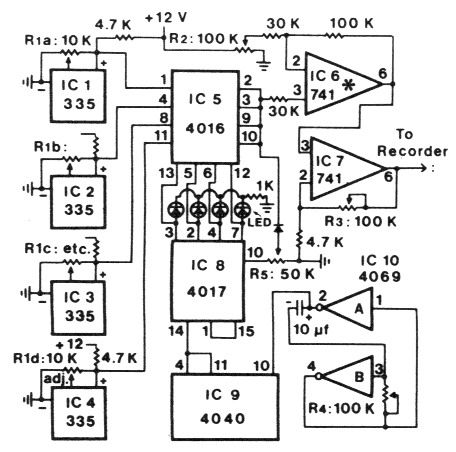
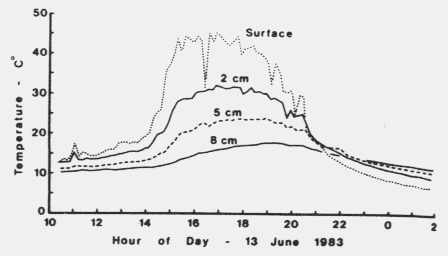
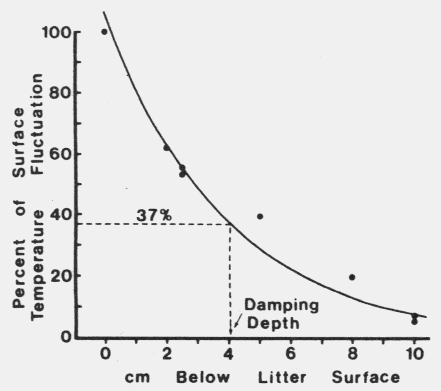
| Table 1. Daily temperature fluctuations at several depths in Norway spruce foret litter during three deys in June 1983 in southern Norway | |||
| Daily temp. fluctuations (oC) | |||
|---|---|---|---|
| June | |||
| Litter depth | 10-11 | 12-13 | 13-14 |
| Sufrace | ±19.6 | ±19.6 | ±19.6 |
| 2 cm | - | - | ±12.2 |
| 2.5 cm | ±10.9 | ±10.4 | - |
| 5 cm | - | - | ±7/6 |
| 8 cm | - | - | ±3.9 |
| 10 cm | ±1.5 | ±1.0 | - |
|
JOHN A. BYERS Department of Ecology, Animal Ecology, Lund University, SE-223 62 Lund, Sweden Present address:
|
|---|